

菌根(Mycorrhiza)是自然界中普遍存在的高等植物与微生物共生的现象。1885年德国植物生理学家Frank首次用Mycorrhiza来描述一些树种的根与真菌形成的联合体,至今菌根研究已有120多年的历史。根据共生体结构特征的明显不同,菌根主要分为两种类型:外生菌根(Ectomycorrhiza)和内生菌根(Endomycorrhiza)。此外,还有一些特殊的菌根类型,如兰科菌根(Orchid Mycorrhiza)和杜鹃科菌根(Ericoid Mycorrhiza)。外生菌根主要存在于树木根系,在植树造林中具有重要作用,而近85%的植物科属和几乎所有的农作物能够形成内生菌根,其在自然生态系统和农业生产实践中的重要作用也日益广为人们所知(李晓林和冯固,2001;刘润进和陈应龙,2008; Smith and Read, 2008)。泡囊-丛枝菌根(Vesicular- Arbuscular Mycorrhiza,VAM)是宿主范围最广、与农业生产关系最为密切的一类内生菌根真菌,因其在植物根内产生“泡囊”(Vesicles) 和“丛枝” (Arbuscules) 两种典型结构而得名。由于部分VAM真菌不产生根内泡囊,但都能形成丛枝, 故简称丛枝菌根(ArbuscularMycorrhiza,AM)。
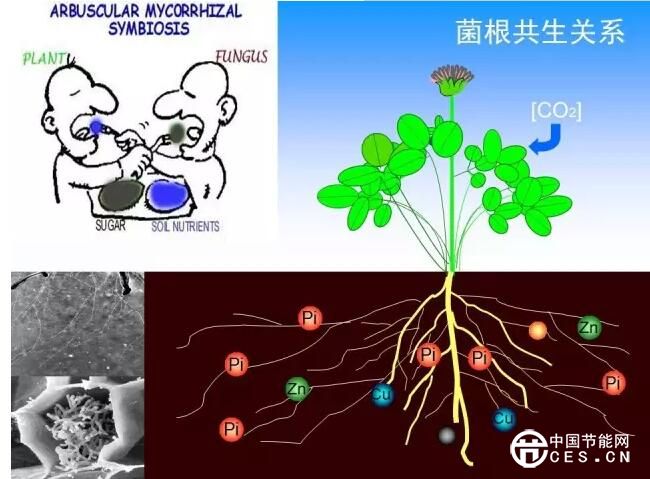
菌根共生体可以通过多种方式或途径影响植物的矿质营养和生长发育过程,在植物逆境生理及群落稳定中有着重要作用。如在养分吸收利用方面,菌根真菌可以通过扩大根系吸收表面积、活化土壤有机磷及难溶性无机磷,及利用菌根真菌对磷的高效吸收运转系统(Ezawa et al., 2002)来提高植物对磷的获取能力。在一些必需矿质养分如铜、锌供应缺乏的情况下,菌根具有类似的作用(Kothari et al., 1991;Li et al., 1991)。对于植物群体而言,菌根真菌可以在时间尺度上稳定植物群落(Koide and Dickie, 2002)。一定程度上,菌根真菌的多样性决定了植物群落的物种多样性、生产力及稳定性(van der Heijdenet al., 1998)。菌根真菌与植物的共生需要植物供应光合产物供应真菌生长,即植物光合产物一部分分配到真菌。菌根根外菌丝体的新陈代谢对于土壤碳库乃至陆地生态系统碳循环具有重要意义(Staddon et al., 2003;Zhuand Miller, 2003)。正是由于菌根真菌具有上述重要作用,其在林业、农作物与经济作物生产中的应用得到广泛深入的研究,而近年来随着生态环境问题的日益突出,人们也开始更多地关注菌根在脆弱或退化生态系统土壤修复中的作用。
2、菌根在植物适应各种逆境胁迫中的作用

菌根共生体系对于宿主植物和菌根真菌适应各种逆境胁迫均具有重要积极意义。在一定程度上,环境胁迫强度也深刻影响着共生双方相互依赖的程度(对于植物来说反映为菌根依赖性)。菌根共生体对逆境胁迫的适应机制一直是菌根研究领域的重要话题,而人们也总是习惯性地更多关注菌根建成对于植物抗逆性的作用,却较少关注逆境胁迫下植物和菌根之间的互动关系,对于菌根真菌自身对逆境胁迫的适应机制更是稀有报道。
(1) 养分贫瘠
菌根真菌能够帮助宿主植物有效获取土壤中的矿质养分,尤其是在土壤中移动性较差的磷及微量元素铜、锌等。增强植物对土壤磷的摄取能力是菌根共生体的最重要功能。菌根真菌对土壤磷的吸收和传输机制是特异高效的,这不仅是菌根共生体系互惠关系的基础,也被认为是菌根帮助植物适应各种逆境胁迫获得更好生长的生理基础。庞大的根外菌丝网络不仅大幅度地延伸了根系吸收范围,而且对土壤理化性质产生影响,从而促进难溶性无机磷的释放;根外菌丝中表达的磷转运蛋白可能直接参与了从土壤中获取磷的过程;菌丝吸收的磷以聚磷酸盐颗粒形式向宿主植物的根部输送;在植物-真菌交换界面-丛枝结构-聚磷酸盐解体释放出磷酸根离子传输给根细胞。总体上,有关菌根真菌吸收并向植物传输磷的生理和分子机制目前已研究得比较系统深入,近年研究热点集中于菌根特异磷转运蛋白的克隆及功能机制,以及不同菌根真菌共生效率差异机制等。类似于一种市场经济现象,植物可能会选择更经济有效的养分吸收途径,并且分配更多的光合产物给贡献更多矿质养分的共生伙伴。
近年来,不少研究者开始关注菌根真菌对氮素的吸收、同化和传输机制,一些研究成果逐步更新了人们对菌根共生体生理功能的认识(Veresoglou et al.,2012)。研究表明,AM真菌根外菌丝可以从周围环境中吸收不同形态的氮素。当NH4+和NO3-并存时,AM真菌更容易吸收同化NH4+。此外,AM真菌还可以吸收利用有机氮,如尿素、Gly、Gln 和Glu等,其中吸收尿素和NH4+比其它氮源速度更快。菌根真菌吸收N后往往是先将N整合入有机N载体-氨基酸,再以氨基氮的形式向植物输送N。在外生菌根中,真菌吸收同化N后产生的自由氨基酸主要是Gln,所以外生菌根以Gln形式向植物运输N。Jin等(2005) 证明了AM真菌在吸收N后储存于Arg,该氨基酸是AM真菌吸收利用N后合成的有机N载体。在根内菌丝体从Arg分解释放出来的N以NH4+形式释放,作为N源可以整合入菌根内的其它氨基酸,也可以传递给寄主植物(Govindarajulu et al. 2005;Chalot et al. 2006)。
外生菌根真菌能够矿化有机质并吸收利用其中的N素,这已基本成为共识。AM真菌也可以通过直接或间接途径调节有机氮的矿化过程。Whiteside等(2009)应用最新的技术-量子点(quantumdots)标记的方法证明AM真菌可以直接吸收、转运有机氮,并且还观察到量子点标记的有机氮能够存在于土壤菌丝、植物根系和植物茎部。AM真菌根外菌丝可能分泌一些与氮矿化有关的酶类,从而加速了有机氮的矿化,然而到目前为止还没有相关直接证据。目前的试验证据主要支持AM真菌通过改变土壤微生物群落结构从而间接加速有机氮的矿化。Atul-Nayyar等(2009)发现AM真菌改变了土壤微生物群落结构,这种变化可能是有机物质迅速降解的原因。
(2) 干旱胁迫
干旱胁迫条件下,菌根共生体依靠其高效的营养物质吸收和转运系统,提高了植物养分吸收效率,缓解了干旱胁迫对宿主植物造成的伤害。然而,AM共生体提高宿主植物抵御干旱胁迫的机制并不仅仅局限于养分的吸收和转运方面。它对宿主植物和生态系统的积极影响还包括提高植物净光合速率;改变进出植物的水流速率,提高根系导水性,增加叶片水势;促进宿主植物某些新陈代谢过程,例如增加生长素合成;影响胁迫响应因子脱落酸(ABA)的合成;改善土壤结构,提高其稳定性等等。尽管目前AM影响宿主植物抗旱性的机制研究相对较多,但总体上还不够系统深入,并且在某些方面还存在争议,而对于菌根真菌增强植物抗旱性的直接作用仍缺乏有力试验证据(李涛等,2012)。最近Li等(2013)从AM真菌Glomusintraradices中克隆了两个水孔蛋白基因GintAQPF1和GintAQPF2,通过酵母异源过表达证明两种水孔蛋白都有明显的输水功能。干旱胁迫条件下,水孔蛋白的高表达导致宿主植物根系相对含水量的显著增加。该研究为丛枝菌根真菌吸收水分提供了直接的分子证据。
(3) 盐分胁迫
盐渍化土壤中存在大量菌根真菌且种类丰富(Wang et al., 2001)。接种AM真菌能促进宿主植物对水分及养分的吸收,提高作物的生长和产量(Sheng et al., 2008)。在盐胁迫下,菌根共生体系能够增强植物对离子的选择性吸收,调节植物抗氧化酶活性和相关基因的表达(Rabie et al., 2005)、改变植株体内的激素水平以及水孔蛋白基因的表达,从而增强宿主植物的耐盐能力。也有研究表明外生菌根通过增强植物细胞质膜的稳定性,提高植物根系的活力,增强植物对水分的利用能力,提高耐盐能力(黄艺等,2006)。
(4) 重金属污染
AM真菌自然具有一定重金属耐性,但重金属污染环境中的菌株其耐性要强于非污染土壤中的菌株(Weissenhorn et al.,1993)。在重金属污染环境中,植物自身获取矿质养分往往比较困难,而菌根能有效增强植物摄取矿质养分的能力,增加其生物量,降低重金属在宿主植物体内的浓度(稀释效应),从而减轻重金属毒害。当然,在此间接作用之外,菌根真菌对植物也有直接的保护效应。AM真菌根外菌丝比表面积大,细胞壁成分如几丁质、 纤维素等对金属具有较强的吸附和吸收能力(陈保冬等,2005),能有效阻滞重金属从植物根部向地上部的转移(Chen et al.,2003;2007);改变土壤pH和根系分泌物的组分从而改变根际微环境,降低重金属对植物的有效性和毒性。目前已发现AM真菌重金属抗性基因,包括抗氧化基因、重金属络合蛋白基因以及重金属转运蛋白基因等(Lanfranco etal., 2002;Waschke et al., 2006;González-Guerreroet al.,2007;Aloui et al., 2009)。
外生菌根发达的外延菌丝能更有效地吸附或吸收重金属离子(Jones et al.,1986),菌根菌套或哈蒂氏网能过滤金属离子(Marschner et al.,1996);菌根分泌的黏液能与重金属结合,阻止其向根部运输(Denny et al., 1987)。目前已从外生菌根中分离到金属硫蛋白基因HcMT1、HcMT2和植物络合素合成酶基因TmelPCS等(Ramesh et al., 2009;Bolchi et al., 2011)。
(5) 有机污染
AM 真菌能直接降解和利用有机污染物, 并将其转化为自身和宿主植物的养分源(刘茵等,2004)。根外菌丝体可以直接富集有机污染物(Verdin, 2005),同时通过增加根系分泌物和特殊性酶的产生(Gramass et al., 1999),以及增强根际其他微生物的活性(Sarand et al., 1998)等途径促进土壤有机污染物的降解。
外生菌根真菌对不同类型的有机污染物都具有一定程度的降解和矿化能力, 在45种已经研究的外生菌根真菌中, 有36种能降解至少一类有机污染物,有12种能降解多种有机污染物(赵曦等,2007)。外生菌根真菌主要通过直接降解、共代谢以及产生多种酶等方式降解有机污染物(Donnelly & Fletcher,1995;Meharg, 2000)。
(6) 病原菌
病原菌可视为植物的非生物胁迫因子。菌根真菌与植物病原微生物都需要从植物获取光合产物,依赖于寄主植物完成生命周期,二者具有类似生态位(niche)。菌根真菌对植物病原微生物的影响主要表现为抑制病原微生物的侵染和繁殖,减轻病害的发生和危害程度,但菌根真菌-病原菌-植物三者之间的相互作用非常复杂,受到多种因素的影响(Hol and Cook,2005;Rafiqi et al., 2012)。菌根真菌提高植物抗病性有直接作用机制和间接作用机制,主要包括:促进植物生长和养分吸收;竞争光合产物及侵染位点(Douds et al., 2000;Deliopoulos et al., 2007);激活植物诱导抗性(Pozo and Azcón-Aguilar, 2007)和改变根际微生物区系(Jaderlun et al., 2008)。其中,菌根真菌诱导植物产生系统性抗性(systemic mycorrhiza-induced resistance)(Cordier et al., 1998; Hao et al., 2012)和对根际土壤中微生物的区系组成和丰度的选择性改变(Aspray et al., 2006; Jaderlund et al., 2008)是当前菌根与病原菌相互作用的研究热点。
3 菌根共生体的生态重要性
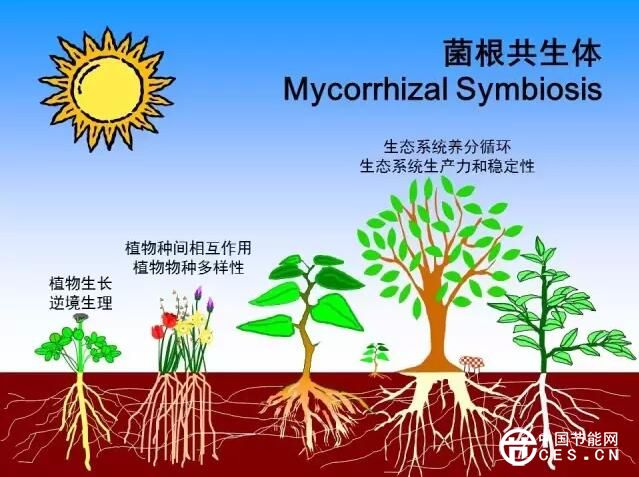
菌根共生体系能够多方面影响植物个体生理和逆境适应性,而在自然生态系统中,菌根真菌能够通过植物根间菌丝桥和地下菌丝网络深刻影响植物种间竞争、群落结构、演替以及生态系统稳定性。众所周知,植物之间除了通过地上部分的不同生长策略以实现空间竞争外,地下部分根系对养分资源的竞争也是其竞争的重要方面。菌根能改善植物养分吸收进而改变菌根化与非菌根化植物之间的竞争关系。特别是在物种入侵方面,菌根真菌可以通过改变竞争作用的强度调节植物群落的组成和外来种的竞争能力,已有研究直接表明AM真菌促进了某些外来种对本地种的竞争优势(Marleret al., 1999)。由于AM真菌与植物共生无严格的专一性,植物群落中植物可通过菌丝桥相互联系传递和分配营养物质,这也是AM真菌对植物群落结构调节重要机制(Read,1997)。在退化生态环境治理方面,应用菌根技术可以加快植被的演替与恢复。
全球气候变化对植物群落结构、演替的影响是生态学家关注的重点,通常认为大气CO2浓度升高条件下,植物的快速生长要求更多的土壤养分,菌根真菌调节植物养分的功能将显得尤为重要和突出。在多种气候因子变化的情况下,探讨菌根真菌对植物群落稳定性的影响将使得我们对生态系统响应气候变化的认识更为客观和全面。大量研究表明,菌根化植物比对照植物光合作用强,因而菌根能增加地上植被的碳固持(Orwin et al., 2011)。菌根真菌菌丝在土壤中的周转时间较快(Staddon et al., 2003),AM真菌的菌丝及其蛋白产物(球囊霉素,Glomalin)能通过促进土壤团聚体的形成和构造而影响土壤有机碳的周转。菌丝通过“黏合-连接-打包”机制将土壤颗粒紧密联系,以此增加有机残片在土壤大团聚体中的存留时间,而球囊霉素作为一种碳蛋白,在土壤中也能起着胶水黏结作用,这一机制将显著提高土壤有机碳,增强土壤碳固持(Rillig and Mummey, 2006; Wilson et al., 2009)。不过,最近的研究表明,在CO2浓度升高的情况下,AM真菌增强了土壤有机碳分解(Cheng et al., 2012),这改变了研究者对菌根增强土壤碳固持的传统认识,也将激发进一步的系统研究。
随着全球变化生物学的兴起,对全球变化和人类活动影响下菌根真菌多样性演变规律的研究也成为菌根生态学的研究热点。运用微生物生态学和生物地理学的原理和方法,从宏观区域尺度上考查菌根真菌在不同环境条件下的群落组成及多样性演变的基本特征,探讨菌根真菌多样性变化的生态效应及主控因子,可为保护脆弱生态区中生物多样性以及脆弱生态系统生态重建提供理论支持。值得注意的是,近年来发展起来的高通量测序技术(如454焦磷酸测序)对于推动菌根真菌多样性研究起到重要作用(Öpik et al., 2009; Lin et al., 2012)。与传统的研究方法相比,高通量测序技术快速高效,且更够更为真实地反映菌根真菌多样性及群落结构,预期高通量测序技术将在菌根生态学研究方面得到更为广泛的应用。
4、菌根应用:资源调查与保藏、应用技术
4.1 外生菌根
随着人们对菌根研究的不断深入,外生菌根在稳定生态系统、农林业生产和社会经济等方面所表现出的重要性越来越引起人们的关注。目前外生菌根技术在引种、菌根化育苗造林、逆境造林、植物病害防治以及食用菌生产等方面的应用都已取得了初步成效。与国际外生菌根研究相比我国起步较晚,但发展迅速,至今已在许多研究领域取得重要进展,有些成果已经达到甚至超过世界先进水平。如:外生菌根真菌Pisolithus tinctorius制剂的研发和利用、截根菌根化育苗和造林技术、对主要造林树种的菌根应用技术等等。当前我国外生菌根研究具有向实际应用方向发展的趋势,目前已投资建成菌剂生产线,年产可达1000t,应用面积每年可达数万hm2。食用菌根菌应用研究方面,云南正着重研究干巴菌(Thelephora ganbajun)、松茸(Tricholomamatsutake)和野生绿菇(Russula virescens),四川等省正在开展对块菌(Tuber spp.)的应用研究。
当前我国外生菌根研究与我国菌根资源大国的地位还不相符合,与国际本领域研究进展相比仍显落后(于富强和刘培贵,2002)。主要表现在:(1)基础研究严重滞后。许多重要经济树种的外生菌根共生关系尚不清楚,资源调查力度不够,家底不清,适合于我国主要造林树种和不同立地条件的优良外生菌根真菌研究不够。(2)尚缺乏足够的分离、培养、保藏、扩大繁殖及菌剂研究和生产的成熟技术,仅限于少数种的应用生物技术也没有得到充分的推广应用。(3)对造林树种菌根化育苗和造林的配套技术缺乏系统的研究。目前,我国尚无一树种具有完整的菌根应用配套技术。
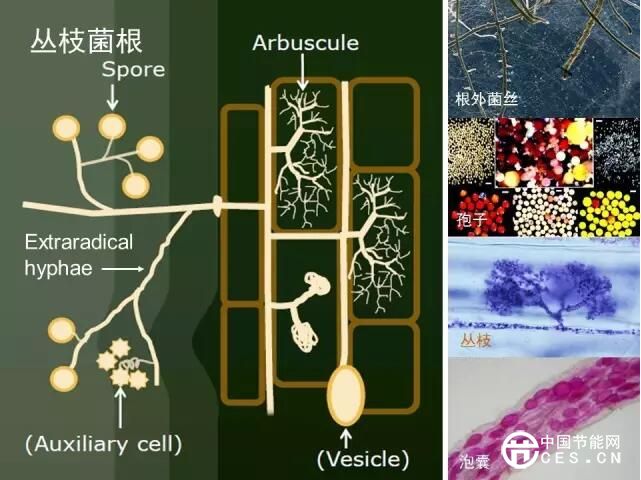
4.2 丛枝菌根(AM)
由于AM真菌的广谱生态适应性和潜在的生态及农业应用价值,国际上一直非常重视AM 真菌的资源调查与分类研究。2010年Schüβler 和 Walker对AM真菌的分类系统进行了总结,归为4目11科18属246种,其中球囊霉科(Glomeraceae) 是种类最多的1个科,下设4属,包括110个种(http://schuessler.userweb.mwn.de/amphylo/amphylo_species.html)。随着分子鉴定方法的发展与应用,出现了很多如Racocetra、Funneliformis、Claroideoglomus等新属,这将大大冲击AMF群落多样性的研究,但分子鉴定方法所带来的研究便利也将极大推动菌根真菌多样性和生物地理学的发展。
由于AM真菌营养方式特殊,只能通过与植物形成互惠共生体才能完成生活史,至今还不能纯培养繁殖。因此,世界各国的AM真菌的资源收集与保藏机构都是和普通、工业、农业及医学微生物菌种库分开而独立建成的。目前,国际性和地区性AM真菌种质资源保藏机构有4个:(1)国际AM真菌保藏中心(International Culture Collection of Arbuscular and Vesicular-Arbuscular Mycorrhizal Fungi,INVAM,http://invam.caf.wvu.edu/);(2)亚洲菌根真菌保藏中心(Centre for Mycorrhizal Culture Collection, CMCC, http://mycorrhizae.org.in);(3)欧洲AM真菌保藏库(European Bank of Glomales,BEG,http://www.kent.ac.uk/bio/beg/)及(4)加拿大与比利时AM真菌根器官培养物保藏中心(Glomalesin vitro collection,GINCO-CANand GINCO-BEL,http://www.mycorrhiza.be)。我国最早最大且拥有自主知识产权的AM真菌种质资源库(Bank ofGlomales in China, BGC)是在北京市农林科学院植物营养与资源研究所建成的,专门从事AM真菌(Glomeromycota)资源收集与保藏工作,现AM 真菌资源保藏总量达到5属25种共136株,已成为国内菌根界独树一帜、影响广泛,专门从事AM真菌种质资源保藏及提供服务的单位(王幼珊等,2012)。
AM真菌纯培养技术还没有突破,目前只能采用盆培养法、静止液培法、流动液培法、喷雾液培法等来生产,成本很高,限制了其大面积的应用。在国际上,有不少国家都有AM菌剂的生产,但生产方式一般还是在温室里用灭菌盆钵培养法以植物为载体生产菌根接种剂。此外也有利用发根农杆菌(Agrobacterium rhizogenes)的Ri质粒转移到胡萝卜的不定根上,使其自身形成生长激素,在人工培养基上迅速生出大量的根,从而加速AM真菌繁殖。目前我国也已基本具备AM真菌菌剂生产技术和高效菌种,但由于缺少基础设施和设备的投入,至今没有进行大规模工厂化生产。
5、展望
在资源和环境问题日益严峻的背景下,菌根技术作为一条有效可行的生物学途径,能够发掘生物的自身潜力,提高植物对自然资源的利用效率,从而降低能耗,减轻环境所承受的压力,必然有着广阔的应用前景。目前,国际菌根研究正日益广泛深入,无论是在宏观生态学方向,还是微观逆境生理方向,都不断取得新的进展。应用方面已经有商业化接种剂生产技术,这为生产实践中大规模应用菌根技术提供了可能。
在菌根技术应用方面,大规模菌剂生产技术及菌根产品(食用菌)商业化将继续发展,在我国可能出现专门的AM菌剂供应公司,可能针对不同的应用领域形成专门化的菌剂生产体系和产品类型;系统的菌根真菌资源调查与保护工作势在必行,对于特殊生境(尤其是极端环境)中的菌根真菌资源的发掘研究与利用需要增加力度,目前的菌根真菌资源保藏体系还需要进一步扩大规模,完善和规范管理及利用模式,注重知识产权保护;对于菌根技术的应用还需要有新的理论和理念,菌根技术的应用要充分考虑适用的条件和范围,外源施加菌根接种剂的理念需要得到拓展和突破,基于生态安全考虑,应考虑将菌根接种技术与根际微生态调控技术结合,充分挖掘利用土著菌根真菌,或是间接调控菌根真菌多样性与丰度,以最小的生态扰动和资源消耗产生最大的生态和经济收益。
我国生态环境复杂多样,由此也产生了高度的生物多样性,包括丰富的菌根真菌资源,这为我国菌根基础研究和应用提供了得天独厚的条件。不过,应当看到目前我国菌根研究和发达国家尚存在明显差距,不仅少有引领国际前沿的研究成果,在菌根真菌资源保护及应用技术方面也相对落后。今后我国的菌根研究者可能要根据我国地域特色、独特的生态环境问题和资源优势,确立菌根研究的生长点与突破口,加强联合研究,实现基础材料和基础理论研究方面的突破,另一方面要积极参与国际交流与合作,提升国际影响力,从而确立我国在国际菌根研究领域应有的地位和作用。







