

厌氧消化技术主要用于处理有机废物,如高浓度有机废水、农业废弃物、餐厨垃圾、剩余污泥等;反应同时产生的生物能源为缓解全球气候变化和保证能源的可持续性提供了新的选择。相比好氧生物处理,厌氧消化具有高有机负荷、剩余污泥量小、绿色能源回收以及较低的运行和维护费用等优势,得到广泛应用并迅速发展。然而,一些环境因素如pH、温度、有机负荷和氨氮浓度(自由氨+铵离子)等影响着厌氧消化过程的稳定进行。其中,氨的毒性抑制被认为是影响厌氧消化过程的最主要因素,自由氨(FAN)起主要抑制作用。研究表明氨产生抑制效果的质量浓度范围是1700~7000mg/L。这一浓度范围受到底物性质、产甲烷菌种、环境因素(温度、pH)以及驯化时期等条件的影响。随着厌氧消化反应器内蛋白质、尿素的分解,氨在消化液中不断累积,逐渐形成的高浓度氨氮将严重影响产甲烷菌的活性,从而降低反应器产气效率,甚至最终导致反应失败。
目前,国内外学者针对厌氧消化的氨抑制进行了大量研究,主要包括氨抑制机理,影响因素以及氨抑制解除方法等方面,然而对于大量恢复方法的研究鲜有系统的归纳总结。笔者通过对近年国内外氨抑制研究的总结,分析和阐释氨抑制的形成机理,从氨浓度和微生物两个角度出发重点归纳了氨抑制的解除方法和技术,并提出进一步发展氨抑制解除技术的研究重点。
1氨抑制的机理研究
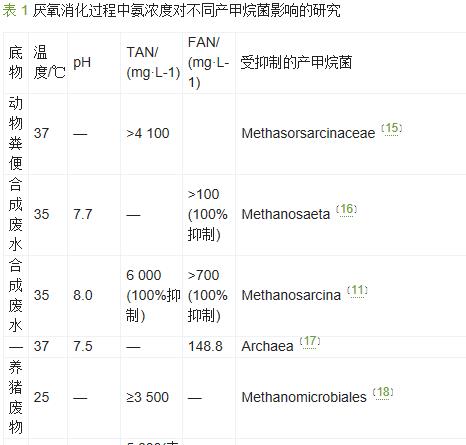
氨是厌氧消化过程中微生物降解蛋白质和尿素等物质的最终产物。在水溶液中,氨主要以离子态氨(NH4+)和自由态氨(NH3)这两种形式存在,其和称为总氨氮(TAN),用以衡量氨的浓度水平。基质中低浓度的氨可以作为微生物的营养物质,而高浓度的氨会严重降低微生物活性。T.Liu等研究认为,TAN的适宜质量浓度为200mg/L。当TAN质量浓度超过3000mg/L时,厌氧消化过程在任何pH条件下都会受到抑制。
目前,对氨抑制机理的研究认识还不完善,已经提出的机理假设有:(1)FAN引起细胞内pH的改变;(2)NH3与K+交换造成微生物细胞内K+的缺乏;(3)额外能量消耗对细胞正常生理活动的影响;(4)NH4+对酶反应的直接影响。G.D.Sprott等在纯甲烷菌种的研究中发现FAN由于其疏水性可直接透过细胞膜进入细菌细胞内,并结合细胞外的H+引起细胞内pH的改变,影响产甲烷菌正常生理活动;同时,NH3在细胞内转化为NH4+引起电荷变化,致使甲烷菌耗能将K+移出细胞以保持电荷平衡,造成细胞内K+的缺乏,进一步降低了产甲烷菌的活性。此外,G.D.Sprott还指出NH4+可能直接作用于甲烷合成过程中的某些酶,导致甲烷的合成失败。W.Wiegant等探究氨抑制机理认为,高浓度TAN严重影响甲烷菌利用H2合成甲烷的途径;另一些中间产物如丙酸、H2的累积,可能抑制乙酸合成甲烷的途径。
稳定的厌氧消化过程主要依赖于水解发酵菌、产酸菌和产甲烷菌3种微生物的正常生理活动。其中产甲烷菌对氨浓度最为敏感,厌氧消化处理马铃薯汁的实验中发现,TAN质量浓度范围在4051~5734mg/L时,产酸细菌基本未受影响,然而56.5%的产甲烷菌活性丧失。在高温厌氧消化的研究中,R.Borja等研究发现乙酸利用型产甲烷菌对氨更加敏感。然而W.Wiegant等的研究认为高浓度氨更易影响H2和CO2合成甲烷的过程。其原因可能归结于氨浓度高低的影响,当TAN质量浓度超过1700mg/L时,氨对乙酸利用型产甲烷菌的抑制作用强于H利用型甲烷菌,而当低于这个临界浓度时,H利用型甲烷菌更易受到影响。不同类型的产甲烷菌对氨的耐受浓度如表1所示。
2氨抑制解除技术与方法
当前应对氨抑制的解除技术主要通过两个方面来进行,一是降低高浓度氨,尤其是FAN,通过对pH、温度、碳氮比等因素的调节来完成;二是驯化产甲烷菌种群,通过填料固定、元素添加等方式增强微生物对高浓度氨氮的耐受性,从而使厌氧消化可以在氨氮浓度较高的情况下进行。
2.1高浓度氨的消除
2.1.1pH控制
系统的pH不仅对微生物生理活性存在影响,而且与反应体系内FAN浓度的高低直接相关,FAN与pH的关系如式(1)所示,随着pH的升高,FAN浓度逐渐增加(当温度分别为25、35、55℃时,Kα分别为5.64×10-10、1.097×10-9和3.33×10-9)〔25〕。
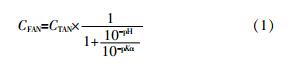
式中:CFAN——FAN的质量浓度,mg/L;
CTAN——TAN的质量浓度,mg/L;
Kα——氨的解离常数。
由较高pH带来的高浓度FAN严重抑制产甲烷菌正常代谢,造成挥发性脂肪酸(VFAs)的累积,降低了系统pH,最终FAN、VFAs和pH相互作用使整个厌氧消化系统达到一种“抑制平衡状态”,这种状态下系统的甲烷产量很低,厌氧消化难以进行。有研究表明氨抑制作用与系统内FAN水平直接相关,而非铵离子的影响。因而可以通过控制pH降低FAN浓度来减轻氨的毒性作用。
P.Shanmugam等〔27〕处理皮革废水,将pH从8.5降低至6.5获得了最佳产气效果,累积沼气产量6518mL。G.Zeeman等〔28〕在TAN为3000mg/L时,将系统pH从7.5调节至7.0,也发现甲烷产量提高了4倍。此外,M.Kayhanian等〔29〕研究高温厌氧消化时发现,pH为7.2,TAN为1000mg/L的条件下系统中对应FAN质量浓度为55mg/L(氨抑制安全阈值);然而pH升高至7.5,TAN仅为400mg/L时,对应FAN浓度便达到这一安全浓度,因而M.Kayhanian建议高温厌氧系统pH应控制在7.0左右。
2.1.2温度控制
温度对微生物的生长速率和FAN浓度均有影响,通常在保证产甲烷菌活性的前提下,厌氧消化工艺选择在中温(30~40℃)和高温(50~60℃)两个范围下进行。由式(1)知FAN浓度与Kα相关,由于Kα与温度成正相关,随温度升高厌氧消化系统内高微生物活性和FAN浓度都会得到提高,因而产甲烷菌的代谢活动有可能受到抑制。研究了FAN浓度与厌氧消化温度和pH的具体关系,结果表明:相同pH条件下,系统反应温度越高,FAN所占TAN比例越大。
研究者发现,高温产甲烷菌相比中温产甲烷菌对氨抑制具有更强的耐受能力。C.Gallert等〔30〕利用厌氧消化处理有机垃圾,中温条件下甲烷产量在氨质量浓度为220mg/L时降低了50%;而高温条件下,氨质量浓度为690mg/L时才引起甲烷产量50%的下降幅度。然而,就整个系统而言,高温厌氧消化过程相对中温厌氧消化过程更容易受到FAN抑制作用。A.G.Hashimoto等〔31〕将氨质量浓度逐步提高超过3000mg/L时,高温消化系统难以维持,而中温系统能在4000mg/L的条件下继续运行。此外,I.Angelidaki等〔32〕在40~64℃范围内研究认为,当FAN质量浓度超过700mg/L时,为保证系统不受氨抑制应当控制反应温度在55℃以下。
2.1.3调节碳氮比
过高的碳氮比会引起系统氮源的不足,无法充分消耗碳源;低碳氮比又可能造成氨的积累而抑制厌氧消化,因而选择合适的碳氮比对厌氧反应器消除氨抑制作用及其稳定运行至关重要。M.Kayhanian等〔29〕对一定范围碳氮比(8~125)下的厌氧消化反应进行了研究,结果发现控制碳氮比在27~32时最有利于阻止氨抑制现象发生和保持产气稳定。在此基础上,O.P.Karthikeyan等〔33〕进一步研究了两种碳氮比(27和32)下的氨抑制作用,结果表明,当碳氮比为32时,系统中的氨浓度相比碳氮比为27时减少了30%。
碳氮比的调节一般可通过不同反应底物间的混合来完成,相比其他方法,具有经济合理、易于操作和增加产气量等优点。P.Shanmugam等〔27〕利用城市生活垃圾与皮革废水混合调节碳氮比为15时,得到最大产气量并控制了氨抑制。但是,碳氮比的调节过程相对较为缓慢,必须在系统被完全抑制之前进行,如果氨抑制作用下系统已经出现VFA积累、pH下降等现象,即使调节至合适的碳氮比,系统也难以从抑制状态恢复〔29〕。
2.2微生物的强化
2.2.1产甲烷菌驯化
驯化接种是增强产甲烷菌氨适应性的有效途径之一。随着系统内氨浓度缓慢增加,微生物可以逐渐适应较高氨浓度的环境。通常,TAN质量浓度为3000mg/L时可完全抑制产甲烷菌,但经过驯化的产甲烷菌可以在高于3000mg/L的环境中生存。I.Koster等〔12〕实验发现驯化后的厌氧消化系统在质量浓度高达11800mg/L的环境下仍然能产生甲烷。而A.G.Hashimoto等〔31〕对比未驯化和驯化的厌氧系统,发现引起氨抑制时的临界TAN质量浓度分别为2400、4000mg/L。
很多研究者都观察到经驯化的产甲烷菌对氨的耐受性明显提高,但是对这种抗性的产生机理还未达成共识,可能是产甲烷菌改变了自身合成甲烷的途径或是新的高抗性产甲烷菌种的出现。F.Lü等〔34〕在研究酸和氨协同作用对产甲烷途径改变的影响时发现,随乙酸含量和氨浓度升高,甲烷合成途径开始由乙酸利用型产甲烷菌降解乙酸途径向由互养型乙酸氧化(SAO)细菌与H2利用型甲烷菌的共生菌团降解乙酸途径转化;当TAN超过6000mg/L,甲烷完全由共生降解途径合成。然而,I.A.Fotidis等〔19〕在类似的研究中却发现了相反的转化,在经过高浓度氨和乙酸环境中驯化后的系统中提高氨浓度,结果产甲烷途径由SAO细菌与H2利用型产甲烷菌(Methanobacterialesspp.和Methanomicrobialesspp.)共生途径转变为乙酸利用途径(Methanosarcinaceaespp.);未驯化的系统暴露于7000mg/L环境时,未观察到产甲烷的途径发生转变,而Methanosarcinaceaespp.为主要的产甲烷菌。
最近的研究中,Methanosarcinasp.(甲烷八叠球菌)作为一种具有超强耐受性的产甲烷菌被重新提出〔35〕。Methanosarcinasp.可以利用乙酸或H2两种途径合成甲烷,并且其形成的菌落拥有较大的比表面积,这些特征使Methanosarcinasp.可以在7000mg/L等严酷条件下生存,因而有研究者认为,将SAO细菌与Methanosarcinasp.构成的共生菌团接种于反应器中能够提高厌氧消化过程的稳定性。但是近期一些关于接种SAO共生菌团的研究并未成功〔36〕,可能是由于产甲烷菌生长速率过慢导致。
2.2.2微生物固定
在厌氧反应器内添加不同的惰性材料(黏土、沸石和活性炭等),通过吸附、离子交换、扩大微生物菌落比表面等作用,能够减轻氨抑制、稳定厌氧消化过程〔37〕。C.Tada等〔38〕研究了不同沸石对氨的去除效果,结果发现天然丝光沸石不仅能去除氨,而且在高TAN条件下(4500mg/L)能够促进甲烷产量。K.Sasaki等〔39〕向厌氧消化反应器添加碳纤维织物(CFT),在TAN质量浓度为3000mg/L时,检测到大量的产甲烷古细菌和Methanosarcinasp.附着在CFT上;未装载CFT的反应器则在TAN质量浓度为1500mg/L时即已表现出产气率下降。
2.2.3元素添加
研究表明一些常量元素的离子(如Na+、Mg2+、Ca2+等)可以削弱或消除厌氧消化反应中氨的抑制作用。B.Demirel等〔40〕研究M.hungatei内的K+传递时发现,当M.hungatei受氨作用被抑制后,添加Mg2+或Ca2+使甲烷菌重新恢复活性。另有研究发现,Na+有助于Methanosarcinabarkeri抵抗氨的毒性作用,而Ca2+可以增强Methanothrixconcilii的氨抗性。
某些微量元素(Se、Co等)的添加,也有利于增强微生物对高浓度氨的抵抗能力〔40〕。C.J.Banks等〔41〕通过添加Se和Co(1.6×10-4g/kg和2.2×10-4g/kg),TAN质量浓度即使在5000~6100mg/L的条件下,厌氧消化反应也能稳定进行。
2.3其他方法
通过一些物理化学方法,如稀释、吹脱、化学沉淀等也可以达到去除氨的目的。H.B.Nielsen等〔42〕向稳定的厌氧消化反应器投加氯化铵验证4种不同稀释方法的恢复效果。结果显示,用新鲜牛粪稀释后的厌氧消化反应得到最高的产气速率,牛粪消化液稀释的系统表现出更稳定的恢复过程。M.Walker等〔43〕发现吹脱法可以有效去除消化液中的氨,并且去除效果符合关于TAN的一阶动力学模型;而且实验证明原位吹脱法能够适应较大范围的有机负荷率,消化前氨吹脱法更有利于对各个工艺单元的操作控制。S.Uludag?鄄Demirer等〔44〕厌氧处理牛粪时,向反应器加入MgCl2•6H2O以形成鸟粪石(MgNH4PO4•6H2O)沉淀来去除消化液中的氨,反应如式(2)所示,结果提高了对NH3去除率。
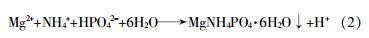
3总结
厌氧消化氨抑制过程复杂,其抑制作用受pH、温度、底物、微生物等多方面因素影响。目前关于氨抑制解除方法的研究主要集中在调节工艺参数控制FAN浓度和通过驯化提高微生物对氨的耐受能力。
然而,通过调节工艺参数(pH、温度、碳氮比等)来控制单一因素——氨浓度,必然会引起其他因素的变化,如降低pH减轻氨抑制的同时却增加了系统VFA的浓度,亦有可能导致厌氧消化过程受VFA累积抑制;降低反应温度可消除氨抑制作用,但同时厌氧消化过程中各类微生物反应活性也会受到影响,造成甲烷产量下降。因此,对工艺参数的调节最终将使厌氧消化反应达到一种相对的平衡状态,甲烷产量达到一定阈值。惰性材料和元素添加则由于经济性问题,难以大规模使用。因而通过微生物驯化增强氨抗性是继续发展解决氨抑制问题的主要途径,未来研究重点可偏向于缩短驯化时间、提纯优良甲烷菌种、稳定接种方式等。







